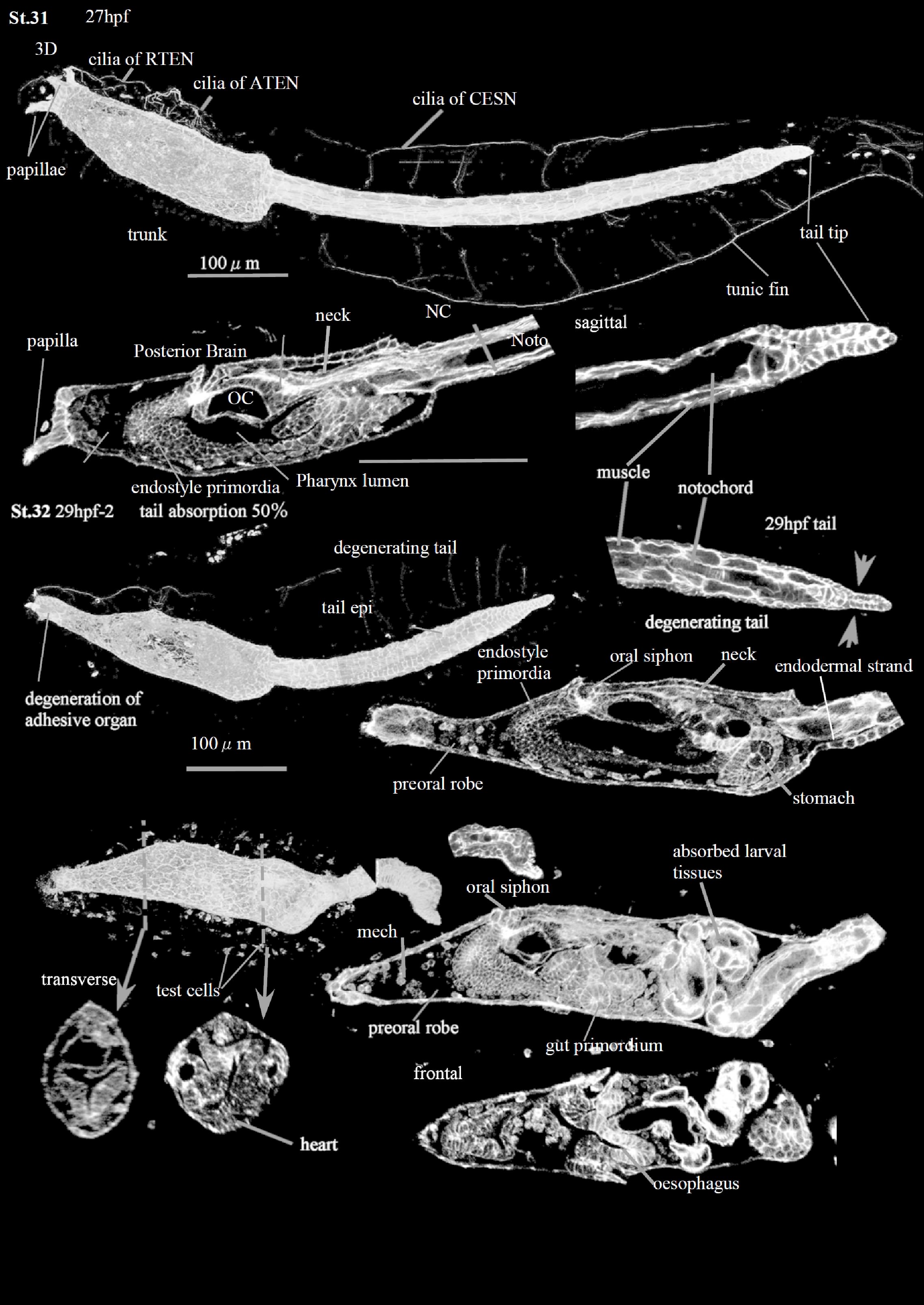
Stage 31
At the Early Tail Absorption Stage (CirobuD:0000054) 27-8 hpf), the shrinkage of the tail epidermis (CirobuA:0000738) begins at the tail tip (Tail tip epidermis: CirobuA:0000949). In the same time, the tail epidermal cells, originally flat, change into thick and cuboidal. The actin staining in the posterior tail is relatively strong, indicating the actin’s involvement in the shrinkage process (Matsunobu and Sasakura, 2015). In addition, the tail inner tissues, such as the notochord, the endodermal strand, and the muscles, begin to be arranged irregularly in the posterior tail region. In some individuals, the tail slightly bends at the trunk-tail transition. The otolith and ocellus are still present, although the larval brain is degenerating; their remnants will be recognizable during the Juvenile Period. The papillae, with their dorsal palp neurons (a8.18 line; CirobuA:0000601) and ventral palp neurons (a8.20 line; CirobuA:0000771), are no longer histologically recognizable at the end of the stage (End stage).
Stage 32
This corresponds to the Mid Tail Absorption Stage (CirobuD:0000055) 28-29 h), when 50% of the tail has been resorbed into the larval trunk. The tail is shorter and thicker than in the previous stage . The tail muscles contract together with the notochord; they fold and stack into the posterior trunk. On the other hand, the tail epidermis (CirobuA:0000738) contracts without folding and finally invaginates into the trunk region. The process recalls the one described during the absorbing of the tail of Halocynthia roretzi and Botryllus schlosseri (Cloney, 1972; Numakunai, 1977). Although the contribution of apoptosis is not excluded (Karaiskou et al., 2015), it has been suggested that the tail epidermis and the extracellular notochord sheath (CirobuA:0000946) generate the strong forces retracting the axial organs into the trunk (Numakunai, 1977). In fact, the tail absorption is inhibited by cytochalasin B, indicating that actin fibers play an important role in the process (Lash et al., 1973).
Concomitantly with tail absorption, the ASNET lose their organization (Terakubo et al., 2010).
The gut continues its differentiation. The oral siphon (osp in S12DI, FI; CirobuA:0000668) opens. The oesophagus (CirobuA:0000621) (Hirano and Nishida, 1997; Nakazawa et al., 2013), the stomach (CirobuA:0000737) (Chiba et al., 2004; Hirano and Nishida, 2000; Nakazawa et al., 2013), and the intestine (CirobuA:0000635) (Nakazawa et al., 2013) are histologically well recognizable. Also, the heart (CirobuA:0000628) is visible (Burighel et al., 1997; Davidson, 2007; Davidson and Levine, 2003; Stolfi et al., 2010; Wang et al., 2013).
The test cells (CirobuA:0000915) are no longer present. They were originally encased in superficial depressions of the developing oocyte by the vitelline coat (Burighel et al., 1997; Chiba et al., 2004; Kawamura et al., 2011; Shirae-Kurabayashi et al., 2006). After fertilization, they were moved into the perivitelline space, to attach to the outer cuticular layer of the tunic. At Stage 32, they are eliminated together with the outer tunic compartment layer and the outer cuticular layer. The stalk starts to elongate.